
I. INTRODUCCIÓN
En los países tropicales, existe una fuerte demanda de productos de origen animal. Los rumiantes pueden cumplir un importante rol para suplir esta demanda, convirtiéndose en una fuente importante de ingresos para los productores (Stürm et ál., 2007). Infortunadamente, los sistemas de producción ganaderos en estos países tienen serios problemas que los hacen poco competitivos. En este sentido, la escasa disponibilidad de forrajes nativos, el alto contenido de fibra y la baja calidad de los mismos representan, quizá, la principal limitante en estos sistemas de producción, especialmente en regiones con suelos ácidos, de baja fertilidad y con largos períodos de sequía (Tiemann et ál., 2008).
Las plantas leguminosas con alto contenido de proteínas han sido sugeridas como recursos promisorios para contrarrestar las deficiencias nutricionales que caracterizan los forrajes de los países tropicales como Colombia; esta alternativa ha suscitado un interés cada vez mayor por integrar estos recursos a la dieta de los rumiantes (Hess et ál., 2006), dado el crecimiento rápido de estas plantas, el alto contenido de proteínas, la buena tolerancia por parte de los animales y los beneficios potenciales, en general, sobre la dieta y la salud de los animales. Adicionalmente, son fijadoras de nitrógeno (N), lo cual reduce o elimina la necesidad de usar fertilizantes nitrogenados en estas regiones. Infortunadamente, muchas plantas leguminosas tropicales se caracterizan por sus altas concentraciones de taninos condensados (TC) que tienen alta afinidad por las proteínas, carbohidratos y otros constituyentes de las plantas, afectando notablemente la digestibilidad de los nutrientes (Waghorn, 2007, Tiemann et ál., 2008). Sin embargo, los efectos negativos sobre la digestibilidad dependen de la concentración de taninos en las plantas, que no guarda un patrón común en éstas. Concentraciones entre 2% y 4% de materia seca son consideradas óptimas para la obtención de beneficios de estas plantas.
¿QUÉ SON LOS TANINOS?
Los compuestos polifenólicos son sustancias ampliamente distribuidas en las plantas, estando a menudo presentes en la dieta de los herbívoros. De estos compuestos, los dos principales grupos polifenólicos, aparte de la lignina, son los taninos condensados y los taninos hidrolizables, los cuales pueden tener efectos tóxicos y antinutricionales o benéficos en el animal (Lowry et ál., 1996, citado por Vitti et al., 2005; McSweeney et ál., 2001). Los taninos se han empleado para preservar la piel de animales destinada a la producción de cuero y para dar astringencia a cierto tipo de bebidas, como el té y el vino (Waghorn, 2007). Conviene anotar que la astringencia es la medida de capacidad para unirse y precipitar proteínas o la capacidad para afectar cambios en la actividad enzimática.
Los taninos de plantas son compuestos fenólicos de naturaleza compleja presentes en una gran variedad de plantas en altas concentraciones y se caracterizan por su capacidad para reaccionar con macromoléculas y proteínas solubles de forrajes durante el paso a través del rumen (Bueno et ál., 2008; Kariuki y Norton, 2008), dependiendo de su estructura química y de su peso molecular. Se unen a proteínas de diferente origen, como plantas, saliva, tejidos, enzimas de los huéspedes y microbios del rumen e intestino (Animut et ál., 2007). Pertenecen a los flavonoides, lo que significa que hacen parte de una de las clases más importantes nutricionalmente, entre las que se incluyen las proantocianidinas, las antocianinas y las isoflavonas. Los taninos son moléculas de alto peso molecular y tienen propiedades para precipitar sustancias como proteínas, alcaloides, ácidos nucleicos, esteroides, saponinas y carbohidratos.
Los taninos se encuentran extensamente distribuidos en las plantas dicotiledóneas, en especial en las leguminosas, capaces de afectar el comportamiento animal; también se encuentran en frutas, árboles y en otras especies, como el sorgo y el maíz, empleadas con frecuencia en la alimentación de bovinos (Provenza et ál., 2000; Min et ál., 2003; Otero e Hidalgo, 2004). Aunque su papel en el metabolismo de las plantas es desconocido, recientemente se han planteado hipótesis acerca de su función en los procesos digestivos de los animales rumiantes.
Diversos factores influyen en la cantidad y clase de taninos que una planta puede sintetizar; la genética y especie de la planta, las condiciones climáticas como la temperatura, la fertilidad del suelo, el estado de desarrollo, el cultivo, el corte y el estado de crecimiento de la planta determinan la concentración de taninos en las plantas. En consecuencia, para determinar dietas particulares e identificar los efectos nutricionales de los taninos en los animales, es necesario cuantificar estos factores y conocer la estructura química y la fisiología del animal objeto de estudio (Stürm et ál., 2007; Bueno et ál., 2008).
De acuerdo con los resultados de diversos estudios, se ha demostrado la notable variabilidad de la concentración de taninos en las plantas, siendo mayor en forrajes leguminosos templados que en forrajes tropicales (Norton, 1999).
Según el grupo biosintético al que pertenecen, los taninos se dividen en:
a) Taninos condensados (TC) o proantocianidinas, los cuales son polímeros flavonoides que pueden ser oxidativamente degradados en ácido a antocianidinas;
b) Taninos hidrolizables (TH), que son derivados de los ácidos gálico y elágico;
c) florotaninos, los cuales se encuentran especialmente en las algas pardas (figura 1).
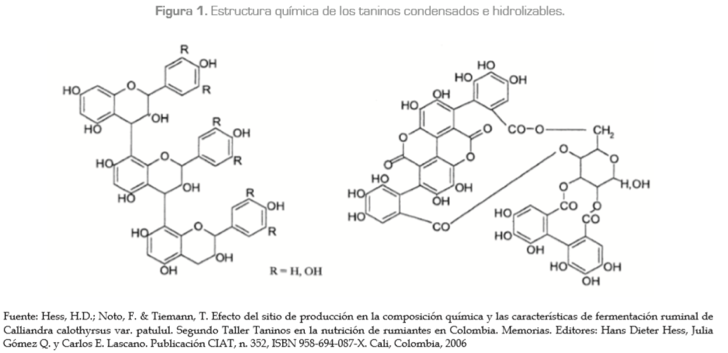
Los TH son moléculas que tienen un carbohidrato en posición central, generalmente de glucosa, cuyos grupos hidroxilos son esterificados con grupos fenólicos como los ácidos gálico o elágico. Presentan bajo peso molecular (500-3.000), son más solubles en agua que los TC y se concentran principalmente en las vainas de las frutas y en las hojas de los árboles. Los productos de degradación de los TH, a diferencia de los taninos condensados, pueden absorberse en el intestino delgado y ser potencialmente tóxicos para estos rumiantes, especialmente cuando se suministran en grandes cantidades sin el suficiente tiempo de adaptación de la flora microbial (Min et ál., 2003).
Los pocos estudios realizados tanto in vitro como in vivo sobre la degradación y con productos de degradación de los TH se han orientado a indagar sobre los efectos tóxicos de éstos en vez de la provisión de nutrientes de los mismos (Waghorn y McNabb, 2003; Rubansa et ál., 2005). Esta situación sugiere que la investigación debe focalizarse en identificar los efectos reales de los taninos en el valor nutritivo y los factores que influyen en sus concentraciones (Romero et ál., 2000). Estos compuestos, además, se degradan rápidamente en grupos fenólicos pequeños, sin capacidad para unirse con las proteínas posteriormente.
Los TC o proantocianidinas, por el contrario, son más comunes y se encuentran difundidos en leguminosas, árboles y arbustos. Son estructuralmente complejos de oligómeros y polímeros de unidades flavonoides (flavan-3-ol, flavan-3,4-diol y biflavanes) unidos mediante enlace carbono-carbono, y no son susceptibles de degradación enzimática anaeróbica. Normalmente se encuentran en las vacuolas de las células y en las paredes celulares de las plantas (figura 2). Estos taninos interactúan bastante en la nutrición de los rumiantes debido a su alta capacidad para ligarse a las proteínas de los forrajes en el rumen luego de la masticación (Andrabi et al., 2005; Stürn et ál., 2007) y a su capacidad para reducir la degradación de proteínas y mejorar su comportamiento cuando las concentraciones de proteína cruda (PC) de la dieta exceden los requerimientos (Provenza et ál., 2000). Sin embargo, cuando las concentraciones de PC en la dieta son bajas y las concentraciones de fibra son altas, los taninos pueden fácilmente tener efectos perjudiciales (Rojas et ál., 2006).
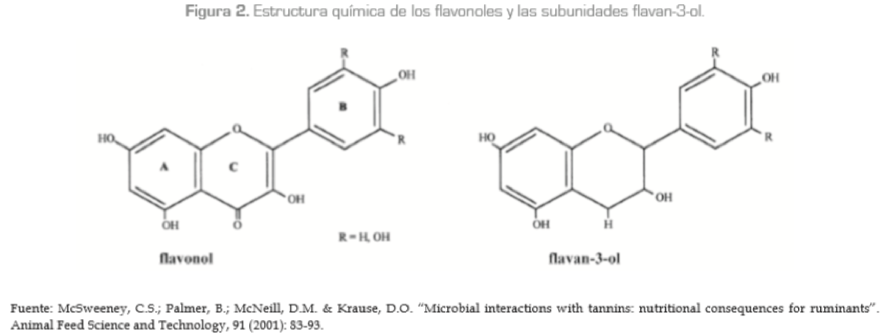
Diversas hipótesis (Vicari y Bazely, 1993, citados por McSweeney et ál., 2007; Waghorn, 2007) señalan que los TC son empleados por las plantas como mecanismo de defensa contra herbívoros y contra patógenos, para la conservación de la energía (para la movilización en caso de necesidad) y para la conservación del nitrógeno, particularmente en ecosistemas de baja fertilidad, pues los TC incrementan siempre el nitrógeno contenido en las heces y reducen la excreción del nitrógeno urinario (Waghorn y McNabb, 2003).
 En general, existen evidencias de que los TC mejoran la ganancia de peso, la producción de lana y la eficiencia reproductiva en rumiantes alimentados con
forrajes de regiones templadas y reducen el impacto del parasitismo gastrointestinal (Nguyen et ál., 2005; Alonso, et ál., 2008). Estos impactos han
orientado las investigaciones a definir la estructura química de los TC, identificar las rutas sintéticas y, especialmente, entender la regulación de la
expresión de los TC en los follajes (Warghorn, 2007). Sin embargo, la respuesta de los animales a la ingestión de TC depende de la concentración de éstos
en las plantas, pues plantas con concentraciones de TC entre el 5% y 10% de la MS reducen el consumo y la digestibilidad del forraje, mientras que
concentraciones comprendidas entre 2% y 4% de la MS favorecen la absorción intestinal de las proteínas debido a la disminución de la protéolisis por parte
de la microflora ruminal (Otero e Hidalgo, 2004).
En general, existen evidencias de que los TC mejoran la ganancia de peso, la producción de lana y la eficiencia reproductiva en rumiantes alimentados con
forrajes de regiones templadas y reducen el impacto del parasitismo gastrointestinal (Nguyen et ál., 2005; Alonso, et ál., 2008). Estos impactos han
orientado las investigaciones a definir la estructura química de los TC, identificar las rutas sintéticas y, especialmente, entender la regulación de la
expresión de los TC en los follajes (Warghorn, 2007). Sin embargo, la respuesta de los animales a la ingestión de TC depende de la concentración de éstos
en las plantas, pues plantas con concentraciones de TC entre el 5% y 10% de la MS reducen el consumo y la digestibilidad del forraje, mientras que
concentraciones comprendidas entre 2% y 4% de la MS favorecen la absorción intestinal de las proteínas debido a la disminución de la protéolisis por parte
de la microflora ruminal (Otero e Hidalgo, 2004).
Tiemann et ál. (2006) evaluaron in vitro el efecto de diferentes concentraciones de taninos provenientes de cuatro leguminosas tropicales sobre la dinámica de la fermentación ruminal; observaron que los TC de Calliandra calothyrsus tuvieron un efecto más negativo sobre la fermentación que los de Flemingia macrophylla y Leucaena leucocephala, postulando que las diferencias de los efectos podían explicarse a partir de la composición química de los TC en estas leguminosas.
Ahora bien, se sabe que el polietilen glycol (PEG) se une a los taninos, posibilitando el incremento de la disponibilidad de los macronutrientes; también se ha observado que animales de diferentes especies, como ovinos, caprinos y bovinos, incrementan el consumo de plantas con taninos cuando son suplementados con PEG (Prichard et ál., 1988, y Titus et al., 1999, citados por Provenza et ál., 2008). Debido a la fuerte respuesta al suplemento de PEG, los animales pueden regular su consumo cuando se les suministran dietas con altos contenidos de taninos, pues se especula que los animales pueden consumir alimentos que atenúen o contrarresten los efectos adversos de los alimentos con taninos.
Para corroborar lo anterior, Provenza et al. (2008) condujeron dos experimentos para indagar, por una parte, si la cantidad de PEG afectaba el consumo de una planta (quebracho) que contenía 20% de taninos y, por otra, averiguar si los corderos se regulaban en el consumo de PEG cuando comían una ración que contenía 0, 5, 10, 15 ó 20% de tanino. Los resultados demostraron que: a) existió una relación positiva entre la cantidad de PEG ingerido y el consumo de planta y tanino (P= 0,0001); b) los corderos que consumieron una dieta alta en taninos, consumieron más PEG que el grupo control (P= .03); c) los corderos que consumieron una dieta con 20% de tanino, consumieron más PEG que los del grupo control; d) el tanino limitó el consumo de la dieta, pero el PEG atenuó la respuesta a un mayor grado (P= 0,065); e) tan pronto se les quitó la ración con tanino, los animales que primero habían consumido raciones con 20% de tanino consumieron más PEG que los corderos de otras raciones (P= 0,0075).
Una de las plantas que más efectos tóxicos ha exhibido en los animales es la Acacia angustissima, una leguminosa tropical originaria de América Central, no obstante sus potencialidades como suplemento proteínico para mejorar la nutricional ruminal.
II. REACCIÓN DE LOS TANINOS CONDENSADOS CON LAS PROTEÍNAS Y OTRAS MOLÉCULAS
Los efectos de los TC sobre los procesos de la digestión están relacionados con la capacidad que tienen para unirse a diferentes moléculas; esta unión depende de la estructura química de los taninos y de diferentes moléculas como celulosa, enzimas digestivas, proteínas, polisacáridos y membranas de bacterias y hongos.
En relación con las interacciones entre los TC y las proteínas, se mencionan cuatro tipos de enlaces: covalentes, iónicos, puentes de hidrógenos e interacciones hidrofóbicas, de los cuales los más frecuentes son los dos últimos, informándose, además, que los enlaces son reversibles y pH dependientes (Min et al., 2003). Así, cada proteína tiene su pH distintivo y óptimo para la unirse a los TC, como la albúmina del suero bovino que forma enlaces con los TC en pH de 4,1, y en pH de 6,1 los TC se unen a la fracción 1 de las proteínas, reportándose que los TC también pueden unirse con proteínas en pH cercano a la neutralidad (3,57,5) y disociarse y liberar a éstas cuando el pH es menor a 3,5 (Jones y Mangan, 1977, citados por Min et ál., 2003).
Otros factores que influyen en el grado de enlaces TC-proteína son el peso molecular, la estructura y la concentración de los TC, así como la compatibilidad de los sitios de unión de las moléculas TC-proteína, el punto isoeléctrico y la configuración de las proteínas. Ahora bien, el significado de estas interacciones en los procesos digestivos para la formación de complejos TC-proteínas es que éstos se llevan a cabo en el rumen con pH entre 5,5-7,2, situación que hace indigestibles a las proteínas por parte de la microflora ruminal, siendo solubles en el abomaso y en el intestino delgado con pH < 3,5 y >8, respectivamente (Andrabi et ál., 2005), lográndose la liberación de las proteínas para que sean digeridas y absorbidas por parte del animal (Hervás et ál., 2003; Min et ál., 2005, McSweeney et ál., 2008).
 Las enzimas son otras moléculas sobre las cuales los TC se unen produciendo efectos inhibidores sobre la actividad de éstas, en particular sobre las
siguientes enzimas: celulasa, ureasa, alfa-amilasa, proteasas y beta-glucosidasa, señalándose que la reducción de la actividad celulolítica puede obedecer
a la disminución de la digestibilidad de la materia orgánica en el rumen. Asimismo, se reporta que las concentraciones de amonio en el rumen se reducen en
virtud de la inhibición de las enzimas proteolíticas que reducen, consecuentemente, la degradación de la proteína de la dieta (McSweeney et ál., 2001; Min
et ál., 2005).
Las enzimas son otras moléculas sobre las cuales los TC se unen produciendo efectos inhibidores sobre la actividad de éstas, en particular sobre las
siguientes enzimas: celulasa, ureasa, alfa-amilasa, proteasas y beta-glucosidasa, señalándose que la reducción de la actividad celulolítica puede obedecer
a la disminución de la digestibilidad de la materia orgánica en el rumen. Asimismo, se reporta que las concentraciones de amonio en el rumen se reducen en
virtud de la inhibición de las enzimas proteolíticas que reducen, consecuentemente, la degradación de la proteína de la dieta (McSweeney et ál., 2001; Min
et ál., 2005).
Otra característica de los TC es la capacidad para ligarse a membranas y a la pared celular de hongos y bacterias, inhibiendo el crecimiento de las bacterias y, por lo tanto, conducen a la reducción de la fermentación ruminal. Adicionalmente, los TC forman enlaces con los carbohidratos, como la celulosa, la hemicelulosa, el almidón y la pectina, informándose que los enlaces con estas moléculas se producen sin relación alguna con el pH.
III. EFECTOS NUTRICIONALES
 La capacidad de los TC para unirse a otras moléculas constituye el aspecto más importante para comprender los efectos que se les atribuyen sobre los
procesos de digestión. Debido a su estructura química poseen la capacidad de unirse a diferentes compuestos como proteínas, polisacáridos, minerales,
carbohidratos, celulosa, células de las membranas bacterianas y enzimas involucradas en la digestión de los compuestos antes mencionados. Durante estos
procesos, los taninos pueden tener efectos positivos y negativos sobre el valor nutritivo de los forrajes según la concentración en la que se encuentren.
Así, a altas concentraciones, 6-10% de la MS deprimen el consumo voluntario y la palatabilidad de las especies forrajeras. También reducen la
digestibilidad: de la materia seca, de la materia orgánica, de la fibra, de la proteína, y de los carbohidratos y por consiguiente afectan negativamente el
desempeño productivo de los animales (Barry y col 1986; Reed y col 1990). En moderada y baja concentración, (2-4 % de la MS), su efecto es beneficioso,
previenen infecciones y aumentan la distribución de nitrógeno no amónico y de los aminoácidos esenciales desde el rumen (Barry y Manley 1984; Mc Nabb y col
1993). La concentración de los taninos en la dieta, con un rango de valores medidos que variaron entre 0 - 12 % de MS, presentaron respuesta lineal y
positiva en la formación de complejos taninosproteínas (Barry y MacNabb 1999).
La capacidad de los TC para unirse a otras moléculas constituye el aspecto más importante para comprender los efectos que se les atribuyen sobre los
procesos de digestión. Debido a su estructura química poseen la capacidad de unirse a diferentes compuestos como proteínas, polisacáridos, minerales,
carbohidratos, celulosa, células de las membranas bacterianas y enzimas involucradas en la digestión de los compuestos antes mencionados. Durante estos
procesos, los taninos pueden tener efectos positivos y negativos sobre el valor nutritivo de los forrajes según la concentración en la que se encuentren.
Así, a altas concentraciones, 6-10% de la MS deprimen el consumo voluntario y la palatabilidad de las especies forrajeras. También reducen la
digestibilidad: de la materia seca, de la materia orgánica, de la fibra, de la proteína, y de los carbohidratos y por consiguiente afectan negativamente el
desempeño productivo de los animales (Barry y col 1986; Reed y col 1990). En moderada y baja concentración, (2-4 % de la MS), su efecto es beneficioso,
previenen infecciones y aumentan la distribución de nitrógeno no amónico y de los aminoácidos esenciales desde el rumen (Barry y Manley 1984; Mc Nabb y col
1993). La concentración de los taninos en la dieta, con un rango de valores medidos que variaron entre 0 - 12 % de MS, presentaron respuesta lineal y
positiva en la formación de complejos taninosproteínas (Barry y MacNabb 1999).
IV. ACCIONES DE LOS TC SOBRE LOS PROCESOS DIGESTIVOS
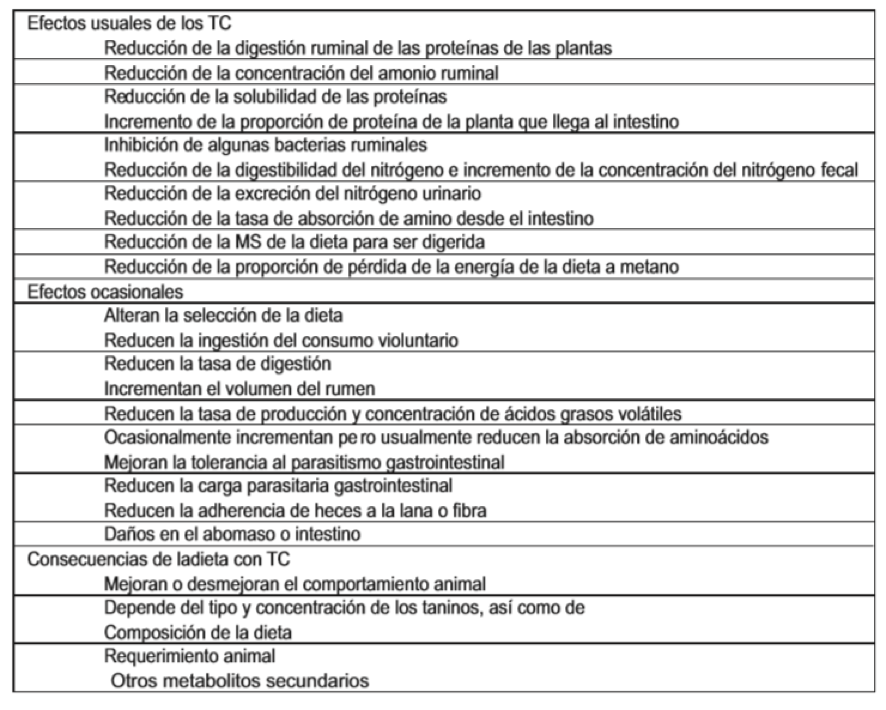
No obstante los resultados contradictorios en relación con los efectos benéficos y perjudiciales de los TC sobre la digestión, existe consenso entre los investigadores sobre los principales efectos de los TC sobre la digestión de los animales, particularmente en rumiantes, y dentro de éstos, los ovinos. Si bien son varios los efectos que los TC producen en la digestión (tabla 4), los principales efectos ocurren en la salivación, la fermentación del rumen, la microflora ruminal, la absorción intestinal, la producción de los animales y la salud animal, especialmente los efectos relacionados con las propiedades antihelmínticas que exhiben estos compuestos sobre los nematodos gastrointestinales de los ovinos, especie en la cual se han llevado a cabo la mayor parte de los estudios (Kaplan y Díaz, 2000; Nguyen et ál., 2005; Iqbal et ál., 2007; Cenci et ál., 2007, Alonso et ál., 2008).
TABLA 1. Efectos de TC en la dieta sobre la digestión de rumiantes.
V. EFECTOS SOBRE LA SALIVACIÓN
Durante el proceso de masticación, cerca del 60% de las células de las plantes se rompe, dejando a los TC dispuestos a ligarse con la saliva y con las proteínas de las plantas, reduciéndose la proporción de proteína soluble en la ingesta y, por tanto, reduciendo la tasa de degradación de proteína. La saliva producida en la masticación contribuye, por un lado, a la formación del bolo alimenticio y a la lubricación del mismo, facilitando la ingestión y, por otro, el bicarbonato y los fosfatos de la saliva cumplen las funciones de buffer para el mantenimiento del pH en el rumen en un rango de 5,8-6,8. En consecuencia, las proteínas de la saliva, especialmente en los rumiantes ramoneadores, mediante la unión con los taninos de la dieta (Waghorn, 2007).
Uno de los órganos productores de abundante saliva son las glándulas parótidas, especialmente en las cabras (especie ramoneadora), que tienen este órgano de gran tamaño en relación con su peso corporal (Vaithiyanathan et ál., 2001), y cuya saliva contiene altas cantidades de proteína rica en prolina (PRP), la cual se une fácilmente a los taninos, dejando menor proporción de TC disponible para unirse a las proteínas en el rumen, siendo ésta la razón principal por la cual las ramoneadoras son más tolerantes a los TC que los ovinos y bovinos.
VI. EFECTOS SOBRE LA FERMENTACIÓN EN EL RUMEN Y LA MICROFLORA INTESTINAL
Los TC afectan la tasa de fermentación en el rumen por la acción que ejercen sobre las bacterias del rumen y sobre las proteínas de las plantas, debido fundamentalmente a la disminución de la protéolisis realizada por las bacterias ruminales y a la unión estable que se da entre los TC y las proteínas. En general, es conocido que la degradación de las proteínas en el rumen ocurre de manera rápida, lo cual implica un uso ineficiente de N para la síntesis de proteína microbial y, por lo tanto, una pérdida de N, debido a que la tasa de degradación de proteínas a amonio es más rápida que la tasa de amonio incorporado a la proteína microbial. En consecuencia, la presencia de TC en la dieta genera una proporción de proteína pasante, no sujeta a degradación en el rumen, pero que posteriormente será sometida a hidrólosis enzimática en el abomaso para ser absorbida en el intestino delgado. Más adelante, los TC forman complejos con las membranas de bacterias y con enzimas extracelulares secretadas por las bacterias, los cuales alteran el crecimiento bacterial y reducen la actividad de las enzimas proteolíticas (Jones et ál., 1993, y McNabb et ál., 1998, citados por Min et ál., 2005).
A los taninos se les identifica como inhibitorios del crecimiento de algunas bacterias del rumen, aunque los mecanismos involucrados no se hayan esclarecido suficientemente. Sin embargo, es probable que las interacciones de los taninos con las bacterias y enzimas producidas por éstas inhiban el transporte de nutrientes en la célula y retarden, por lo tanto, el crecimiento de las bacterias (McSweeney et ál., 2001). Existen diferentes plantas, como el Lotus corniculatus, cuyos TC reducen el número de bacterias del rumen o producen cambios morfológicos, ocasionando una reducción sustancial de la actividad proteolítica de muchas bacterias.
Por ejemplo, Bae et ál. (1993, citado por McSweeney et ál., 2001) reportaron que la actividad de la bacteria Fibrobacter succionegenes y su enzima extracelular endogluconasa fue inhibida por una concentración de 100-400 µg/ml de L. corniculatus bajo condiciones in vitro. Situación similar ocurrió con el crecimiento inhibido de las bacterias proteolíticas Butyrivibrio fibrisolvens, Ruminobacter amylophilus y Streptococcus bovis, por parte de TC (<600 µg/ml) Onobrychris cuneata, aunque la cepa Prevotella ruminícola fue tolerante a esta concentración de TCl, lo cual indica que algunas bacterias proteolíticas son relativamente no afectadas por los taninos (Min et ál., 2005).
Los mecanismos involucrados en la defensa de las bacterias de los taninos son la secreción de un polisacárido extracelular que impide la formación del complejo bacteria-tanino, y la formación de una capa gruesa de glicoproteína que tiene alta afinidad por los taninos (Broker et al, 1994, citado por McSweeney et ál., 2001). Debido al interés manifiesto por identificar los microorganismos tolerantes a los taninos condensados e hidrolizables, se ha logrado aislar cepas que incluyen a S. gallolyticus, cepa muy relacionada con S. bovis, Clostridium sp., provenientes de ovinos, cabras y venados (Broker et al., 2000, citado por McSweeney et ál., 2001).
 En un experimento con ovinos, Min et al. (2003) observaron que cuando cambiaron la dieta de ryegrass/trébol blanco, que no contenía taninos, por una dieta
de L. corniculatus (32 g TC/kg MS), la población de bacterias ruminales proteolíticas Clostriodium proteoclasticum B316, Eubacterium spp. C12b,
Streptococus bovis B315 y Butyrivibrio fibrisolvens C211 decrecieron de 1,6 x 108, 2,7 x 108, 2,7 x 106 y 1,2 x 106 a 5,1 x 107, 1,5 x 108, 1,6 x 106 y 1 x
106 por ml, respectivamente, con lo cual demostraron que la acción de los TC en los forrajes reducen notablemente el crecimiento bacterial proteolítico en
el rumen. Se ha sugerido que la presencia de altas concentraciones de taninos libres pueden interactuar con otro tipo de proteínas, como algunas enzimas
secretadas por las bacterias del rumen, inhibiéndose, porl o tanto, la fermentación de carbohidratos en el rumen (Barry y Manley, 1986, citados por Min et
ál., 2003).
En un experimento con ovinos, Min et al. (2003) observaron que cuando cambiaron la dieta de ryegrass/trébol blanco, que no contenía taninos, por una dieta
de L. corniculatus (32 g TC/kg MS), la población de bacterias ruminales proteolíticas Clostriodium proteoclasticum B316, Eubacterium spp. C12b,
Streptococus bovis B315 y Butyrivibrio fibrisolvens C211 decrecieron de 1,6 x 108, 2,7 x 108, 2,7 x 106 y 1,2 x 106 a 5,1 x 107, 1,5 x 108, 1,6 x 106 y 1 x
106 por ml, respectivamente, con lo cual demostraron que la acción de los TC en los forrajes reducen notablemente el crecimiento bacterial proteolítico en
el rumen. Se ha sugerido que la presencia de altas concentraciones de taninos libres pueden interactuar con otro tipo de proteínas, como algunas enzimas
secretadas por las bacterias del rumen, inhibiéndose, porl o tanto, la fermentación de carbohidratos en el rumen (Barry y Manley, 1986, citados por Min et
ál., 2003).
De otro lado, si bien se conoce poco sobre los efectos de los TC sobre la hidrólisis de la ribulosa-bifosfato carboxilasa/oxigenasa (Rubisco, fracción 1 de la proteína) o acerca de las actividades específicas de miroorganismos del rumen sobre el Rubisco (Min et ál., 2005), Aerts et ál. (1999, citado por Min et ál., 2003) encontraron que los TC extractados de L. corniculatus y de L. pdeunculatus (237477 µg TC/ml) protegieron fuertemente a Rubisco de la degradación por parte de los microorganismos del rumen.
VII. EFECTOS SOBRE LA ABSORCION INTESTINAL
Se acepta que los TC incrementan siempre el paso de la proteína de las plantas al intestino, reduciendo la absorción neta de amoniaco en el rumen, aunque ello no signifique que necesariamente la proteína sea digerida en la porción ácida del abomaso al disociarse de las proteínas en pH por debajo de 3,5. Adicionalmente, cualquier cantidad de TC que no esté ligada a la proteína de las plantas puede ligarse a enzimas endógenas y al lumen intestinal, provocando alteraciones en la absorción intestinal (Waghorn, 2007) de monogástricos, como ratas y cerdos, pero especialmente pollos, como lo demostró el estudio de Ortiz et ál. (1994, citado por Waghorn, 2007) al observar que el suministro de bajas concentraciones de TC en la dieta (1020 g/kgMS) ocasionaba acortamiento de las vellosidades intestinales, distorsión y atrofia de las mismas y proliferación de secreción mucosa, con la consecuente inhibición de la absorción de aminoácidos.
El impacto de los TC sobre la función intestinal en rumiantes no está bien dilucidado, aunque parece que los ovinos son menos sensibles a sus efectos que los monogástricos. Así, por ejemplo, Walton et al. (2001) no observaron cambios en las estructuras de las vellosidades intestinales de ovejas alimentadas durante cuatro semanas con Lotus. Sin embargo, en el estudio de Robins y Broker (2005) se observaron reducción de la actividad enzimática entre un 50% y 70%, daños abomasales y muerte de las criptas abomasales e intestinales al alimentar a ovejas durante cuatro semanas con altas concentraciones de taninos astringentes (Acacia aneura). Estos cambios sugieren que estos compuestos inhiben la capacidad de las enzimas endógenas para desdoblar las proteínas en péptidos y aminoácidos, e inhiben también su absorción, con lo cual se reducen la tasa de hidrólisis de las proteínas y la inhibición de la absorción de aminoácidos. Adicionalmente, la actividad enzimática endógena puede exceder los requerimientos para la protéolisis, pero cuando los TC se unen a la superficie de las bacterias o a las proteínas de las plantas, pueden reducir el acceso y actividad de las enzimas.
Lo anterior indica que los TC son un arma de doble filo a la hora de valorar sus bondades alimenticias, pues ellos no son digeridos por la microflora y el esqueleto carbonado no se absorbe; de esto se infiriere que altas concentraciones de taninos pueden reducir la disponibilidad potencial de MS para la digestión.
Uno de los aspectos más esclarecidos, hasta ahora, es que la respuesta de los rumiantes a dietas que contengan TC depende de su estado fisiológico o productivo (lactantes, jóvenes o adultos); la condición fisiológica afecta sus demandas nutricionales, la calidad de la dieta en su conjunto (fibra, concentración de proteína) y la astringencia de los TC. Así, en varios experimentos se observó que corderos machos jóvenes o lactantes alimentados con leguminosas templadas incrementaron su peso debido a que su organismo respondió favorablemente a las adiciones suplementarias de proteína en la dieta; en cambio no sucedió lo mismo en los animales en los que los aminoácidos no son una limitante para su desempeño (Waghorn, 2007).
VIII. EFECTOS DE LOS TC EN LA PRODUCCIÓN ANIMAL
EFECTOS SOBRE LA PRODUCCIÓN DE LANA
El efecto de los TC sobre la producción y calidad de lana está muy bien documentado y demostrado. Los estudios son concluyentes en el sentido de que el acción de los TC en la producción de lana depende de la concentración y el tipo de TC. Se ha observado que la producción de lana se incrementa en un 10% en animales a los que se les ha suministrado entre 22 y 38 g/kgMS de L. corniculatus; mientras que al suministrar más de 50 g/kgMS de L. pedunculatus la respuesta ha sido negativa. Sin embargo, cuando las con-centraciones fueron inferiores a 22 g/kgMS, la respuesta fue variable, lo cual sugiere que los efectos positivos de L. corniculatus para la producción de lana están en el rango 22-38g/kgMS (Min et ál., 2003).
EFECTOS DE LOS TC SOBRE LA REPRODUCCIÓN
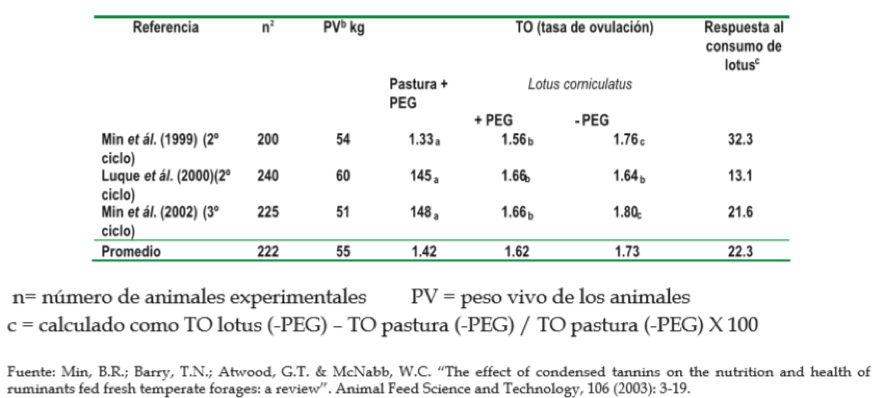
La nutrición de los animales es un factor clave en el desempeño general de los mismos; en particular, es determinante en la reproducción de las ovejas, pues está relacionada con su tasa de ovulación (ver estudios de la tabla 2), aunque los mecanismos involucrados en la tasa de ovulación no estén del todo claros.
Se cree que los efectos de la nutrición sobre la tasa de ovulación estén relacionados con el control metabólico hormonal de los ovarios, y se ha observado cierta correlación existente entre las alteraciones en las concentraciones plasmáticas de la hormona del crecimiento y el factor de crecimiento ligado a la insulina (IGF, por sus siglas en inglés) y las alteraciones en la energía corporal, balance de proteínas y la síntesis de proteína muscular, lo cual puede afectar la función ovárica, sea directamente o mediante la modulación de la acción de las gonadotropinas en los ovarios. Igualmente, se ha detectado que las concentraciones de aminoácidos esenciales y aminoácidos de cadena ramificada están positivamente correlacionados con la tasa de ovulación (varios autores, citados por Min et ál., 2003).
TABLA 2. Efecto del pastoreo sobre L. corniculatus o pastura (Ryegrass perenne/trébol blanco) y suplementación con polietilenglicol (PEG) sobre la tasa de
ovulación en ovejas.
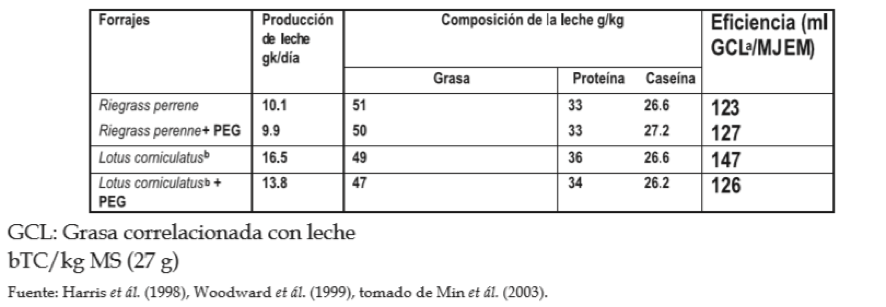
EFECTO DE LOS TC SOBRE LA PRODUCCIÓN Y COMPOSICIÓN DE LA LECHE
Min et ál. (2003) reportan que en un estudio usando vacas de leche alimentadas con L. corniculatus, el incremento en la producción de leche fue de 60% superior a la producción de leche en vacas alimentadas con ryegrass perenne, obteniéndose también incrementos del 10% en la concentración de proteína de la leche. De la misma manera, Wang et ál. (1996, citados por Min et ál., 2003) reportaron incrementos en la producción de leche, de lactosa y proteínas en 21%, 12% y 14% en la mitad y final de la lactación de vacas alimentadas con L. corniculatus, aunque no se observaron incrementos en la fase temprana de la lactancia.
TABLA 3. Efecto de la especie de forraje sobre la producción y composición de la leche en vacas Holstein Friesian.
IX. EFECTOS DE LOS TC SOBRE LA SALUD ANIMAL
Los rumiantes en pastoreo están expuestos a diferentes enfermedades, algunas relacionadas con componentes metabólicos y nutricionales. Dos de las principales afecciones son las enfermedades parasitarias, particularmente las gastrointestinales, y la formación de gas en el rumen, que puede conducir a situaciones de timpanismo.
La formación de gas es ocasionada por la alta solubilidad de las proteínas en el rumen que conduce a la formación de espuma en el rumen, siendo bastante prevalente en bovinos que pastorean leguminosas. Se ha comprobado que los animales que consumen leguminosas que contienen TC reducen notablemente la producción de gas mediante la precipitación de la espuma de las proteínas de las plantas, previendo la aparición de timpanismo por la acción de los TC (Min et ál., 2005; Tavendale et ál, 2005; Animut et al., 2007).
En relación con el parasitismo intestinal, se sabe que las infecciones por endoparásitos ocasionan pérdida de proteínas, lo que se traduce en pérdidas económicas en los sistemas de producción de rumiantes. Adicionalmente, la presencia de resistencia a los antihelmínticos en estas especies hace prioritario que se exploren alternativas no químicas de control, para evitar la extensión de este problema a nivel mundial. Una alternativa puede ser el uso de TC, los cuales, de acuerdo con resultados en diferentes países del mundo, han demostrado ser eficaces para el control de los parásitos del abomaso y del intestino delgado, especialmente en ovejas que, hasta ahora, es la especie de rumiantes más explorada en esta temática. En la tabla 7 se muestran los efectos de los TC sobre el parasitismo gastrointestinal en ovinos (Niezen et ál., 1995, 1998, citados por Min et ál., 2003).
El estudio de Iqbal et al., (2007), con pruebas in vivo e in vitro, analizó los efectos directos e indirectos de los TC sobre una cepa de Haemonchus contortus en corderos alimentados con dietas de bajo y alto contenido de taninos (2% y 3% de TC), comparados con un grupo control sin tratamiento. Los interesantes resultados indicaron que en los corderos con alto y bajo contenido de TC la inhibición de la eclosión de huevos dependió de la dosis, mientras que no se observó efecto sobre los adultos de H. contortus. En las pruebas in vivo se observaron menores recuentos de huevos por gramo de materia fecal que en el grupo control.
En este mismo sentido se conocieron recientemente otros reportes, como los de Alonso et al. (2008), Cecnci et ál. (2007), Hoskin et ál. (2000) y Athanadasiou et al. (2001), en los que se demuestra la capacidad de los TC para actuar no sólo indirectamente sino directamente sobre los parásitos intestinales y pulmonares de ovinos y venados, como Trichostrongylus colubriformis, T. axei, Dictyocaulus sp, y Ostertagia circumcincta.
Si bien los TC se consideran responsables de efectos perjudiciales tanto en animales monogástricos como en los rumiantes, éstos últimos pueden también beneficiarse de ellos cuando son empleados controladamente en sus dietas, lo cual se refleja en el incremento en la ganancia de peso vivo, en mayores producciones de leche y lana y en efectos antiparasitarios. Por lo tanto, se puede afirmar que los TC causan efectos positivos y negativos en los rumiantes. La demostración de estos efectos está soportada por una amplia gama de experimentos en campo sobre la química y fisiología digestiva que ha prevalecido en la investigación sobre los TC en las dos últimas décadas. En la tabla 4 se resumen los principales efectos positivos de los TC en los rumiantes.
Tabla 4. Beneficios potenciales de los TC en la dieta sobre la salud animal y la sostenibilidad ambiental.

X. EFECTOS POSITIVOS DE LOS TC
Desde la perspectiva de los efectos positivos de los TC en los organismos y sistemas de producción de animales, sobre todo rumiantes, su uso debe ser considerado de importancia capital, pues con ellos se contrarresta o reduce la protéolisis que normalmente ocurre en el rumen. De acuerdo con lo demostrado en los diferentes resultados disponibles, se ha definido con claridad que para poder lograr efectos benéficos de los TC, éstos deben estar en un rango de concentración entre 2% y 4% de la MS, considerándose el rango óptimo por no afectar el desempeño productivo de los animales ni el consumo de la dieta.
EFECTOS SOBRE LA PROTEÍNA EN EL RUMEN
Esta función de los TC se da especialmente porque: precipita las proteínas de la ingesta, con lo cual se incrementa su paso al intestino delgado para ser absorbidas, protege la fracción 1 de la proteína (Rubisco); aumenta la absorción de los aminoácidos esenciales y disminuye la de los no esenciales; disminuye la digestibilidad del N de la dieta en porcentajes cercanos al 15%, debido a la menor concentración de amonio ruminal, como consecuencia de la menor degradación de las proteínas, e incremento de la absorción intestinal; reduce la fermentación de carbohidratos por la unión que estable la porción de taninos libres con enzimas microbianas inactivas, disminuyendo, por lo tanto, la digestión de los carbohidratos.
EFECTOS SOBRE LA MICROFLORA RUMINAL
Si bien los resultados arrojados demuestran que las respuestas de la microflora ruminal varían dependiendo de las especies que contienen TC, no hay duda del efecto de los TC para inhibir u ocasionar cambios en la población microbiana ruminal, con lo cual se afecta la capacidad hidrolítica de estas bacterias.
EFECTOS SOBRE EL CICLO DE LA ÚREA
Se ha establecido que el complejo TC-proteína reduce la disponibilidad de nitrógeno para las bacterias ruminales, la degradación y desaminación de las proteínas, reduciendo, por lo tanto, el amonio ruminal; lo cual incrementa la eficiencia del reciclado de la úrea en el rumen.
EFECTOS SOBRE LA PRODUCCION Y LA SALUD ANIMAL
Existe suficiente evidencia sobre las bondades de los TC en la dieta de los animales: incrementan y mejoran la producción de lana en los ovinos, aumentan la producción de leche y la ganancia de peso vivo de los animales y previenen el timpanismo en rumiantes al reducir la formación de espuma en el rumen por la precipitación de las proteínas. En la parte sanitaria, está suficientemente demostrada la capacidad antihelmíntica de los TC y en la reducción de las miasis en los ovinos, así como en el incremento de la resiliencia.
Los efectos negativos de los TC se relacionan con la disminución del consumo de alimentos, debido probablemente a la reducción de la palatabilidad ocasionada por los efectos astringentes de estos compuestos en la saliva, lo cual afecta la digestión de los rumiantes generando consecuencias negativas sobre el crecimiento de los animales. Así mismo, los TC producen sensación de llenado del rumen debido a la disminución de la fermentación, y reducen la digestibilidad de las células cuando se unen a carbohidratos estructurales o a las enzimas de las bacterias.
XI. CONCLUSIONES
El uso de TC en la dieta de rumiantes constituye, probablemente, una alternativa sostenible para la alimentación de rumiantes en las regiones tropicales, a la luz de los conocimientos actuales. Los resultados de diferentes investigaciones han demostrado que los TC tienen efectos benéficos en los rumiantes, entre los que se destacan: el incremento en la producción de los animales, particularmente de leche y lana en ovinos; el mejor el desempeño de los animales frente a desafíos parasitarios gracias al incremento de la resiliencia; y reducción del riesgo de presentación de enfermedades como el timpanismo. Estos efectos se logran en virtud de la capacidad que tienen los TC para unirse a las proteínas de la dieta y reducir, de esta manera, la degradación proteica en el rumen, lográndose una proporción mayor de proteína pasante al abomaso.
Sin embargo, el efecto benéfico de los taninos sobre la digestión de rumiantes está asociado a la cantidad de tanino consumido y a la composición química de los mismos. En relación con esta última, se ha evidenciado que es variable aún entre muestras de las mismas plantas, por lo que se deben emprender mayores esfuerzos de investigación con el propósito de esclarecer los efectos reales de los taninos en las diferentes plantas leguminosas de las regiones del trópico.
Copyright © 2016 Pronutron - Designed by PeruDevOps.